凝血系统(凝血因子)是生物体为了止血,使血液凝固的一系列分子的作用系统。与凝血相对,纤溶系统负责溶解和分解已经凝固的凝块(血栓)。由于这两者在许多病理情况下密切相关,因此本文一起介绍它们。
★凝血系统
一期止血
血管的止血
发生出血时,机体通过收缩外周血管止血,同时泄漏到血管周围的血液压缩血管也发挥止血作用。血小板释放的血清素和血栓素A2参与血管收缩。被破坏的组织细胞释放的前列腺素E2刺激局部神经敏感,并向中枢神经系统发送疼痛信号,以引起机体注意受出血影响的区域。对于微不足道的微小损伤或者威胁生命的重大损伤,中枢神经系统评估体外情况,并根据出血程度做出“战斗或逃跑”的重要判断来客服外部环境,同时分泌去甲肾上腺素进一步收缩外周血管。同时中枢神经系统还能释放内啡肽阻断疼痛信号。
血小板止血
●受伤组织的细胞会分泌血管性血友病因子。
●von Willebrand factor(冯·维勒布兰德因子)与血小板表面结合。
●损伤部位的胶原蛋白和凝血酶激活血小板和磷脂酶C。
●磷脂酶C将二磷酸磷脂酰肌醇 (PIP2) 降解为三磷酸肌醇 (IP3),IP3作为第二信使促进内质网释放Ca⁺⁺。
●血小板在损伤部位聚集并释放血栓素A2、凝血因子V、二磷酸腺苷 (ADP),它们能进一步激活周围的血小板。
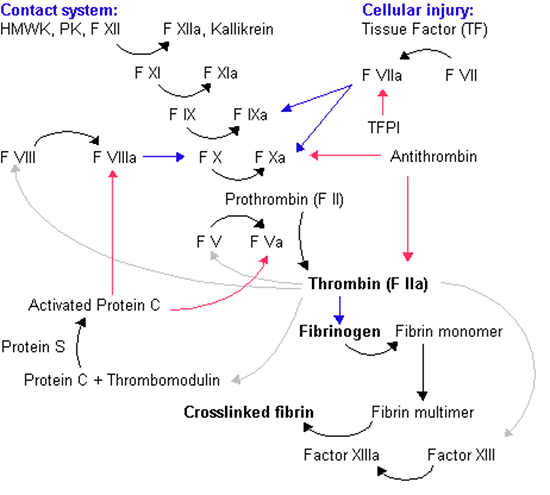
二期止血
开始期
●外源途径
当细胞受损时,组织因子与因子VIIa(被激活的凝血因子VII)结合。
VIIa 激活凝血因子IX,生成凝血因子IXa。
IXa 激活凝血因子X,成为凝血因子Xa。
●内源途径
当血液接触带负电荷的固体(如岩石或沙子)时,前激肽释放酶和高分子量激肽原可激活凝血因子XII,生成凝血因子XIIa。
XIIa激活凝血因子XI,成为凝血因子XIa。
XIa激活凝血因子IX成为因子IXa。
增幅期
凝血酶激活凝血因子XI,生成凝血因子XIa。XIa 激活凝血因子IX,成为凝血因子IXa。
此外,凝血酶本身激活凝血因子V和VIII,分别生成Va和VIII。
进一步,凝血酶激活血小板,以将因子Va 和因子 VIIIa 结合到血小板表面。
传播期
与血小板表面结合的凝血因子VIIIa和IXa激活凝血因子X后,Xa与血小板表面结合。
与血小板表面结合的凝血因子Xa和Va持续将凝血酶原转变为凝血酶。
大量凝血酶将血浆中的纤维蛋白原分解成纤维蛋白单体。大量的纤维蛋白单体被凝血因子XIII交联成纤维蛋白聚合物,并夹带其他血细胞形成血凝块(血栓)。
关于凝血因子
按发现顺序使用罗马数字命名了各凝血因子,而不是通常的自然科学惯例(由发现者命名)。人们不断发现新的凝血因子,有的后来被证明是同一因子的另一种形式。因此凝血因子的数字排列有缺失现象。然而,很少用罗马数字来称呼前四个凝血因子。
●凝血因子I:纤维蛋白原/纤维蛋白
●凝血因子II:凝血酶原/凝血酶
●凝血因子III:组织因子或凝血酶原激酶
●凝血因子IV:钙离子
●凝血因子V (proacceleline):促凝血球蛋白原。一种分子量为33000 的蛋白质,由位于 1 号染色体长臂 (1q23) 的 F5 基因编码,在肝脏中表达并释放到血液中。残基 506 从精氨酸突变为谷氨酰胺的多态型被称为凝血因子V Leiden(来自荷兰城市的名称),并观察到它与静脉血栓栓塞的增加关系密切。在黑色和黄色人种中很少见。此外,由于该基因为常染色体隐性遗传,因此很少见凝血因子V的缺乏症。
●VI是一个缺失的数字,它是凝血因子Va的旧名称(Va指被激活的凝血因子V)。
●凝血因子VII:促凝血酶原激酶原
●凝血因子VIII:血友病因子A。一种分子量约为30,000的蛋白质,由F8基因编码,位于 X染色体长臂 (Xq28) 的末端。在血浆中,它存在于具有冯维勒布兰德因子的复合物中。缺乏它会导致血友病A。
●凝血因子IX:血友病因子B。一种分子量约为55,000至60,000的糖蛋白,由位于X染色体长臂末端(与因子VIII相邻的 Xq27 区域)附近的 F9 基因编码。缺乏它会导致血友病B。
●凝血因子Ⅹ:字体凝血酶原C。一种分子量约为35,000的糖蛋白,由F10基因编码,位于 13号染色体长臂末端附近(与因子VII相邻的13q34 区域),主要位于肝脏中。它以依赖维生素K的方式被合成并释放到血液中。
●凝血因子XI:抗血友病球蛋白C,或血浆促凝血酶原激酶前体。由F11基因编码的80 kDa蛋白,被定位在4号染色体长臂末端附近,形成S-S结合二聚体,与更高分子量的激肽原以1:1结合。
●凝血因子XII:表面因子,或哈格曼因子。
●凝血因子XIII:纤维蛋白稳定因子。
●前激肽释放酶
●高分子量激肽原(菲茨杰拉德因子)
凝血障碍时的检查所见
| 状态 |
凝血酶 原时间 |
活化部分凝血 活酶时间 |
出血时间 | 血小板数 |
|---|---|---|---|---|
| 维生素K缺乏或华法林 | 延长 | 无变化或少许延长 | 无变化 | 无变化 |
| 弥散性血管内凝血 | 延长 | 延长 | 延长 | 减少 |
| 冯维勒布兰德氏病 | 无变化 | 延长或无变化 | 延长 | 无变化 |
| 血友病 | 无变化 | 延长 | 无变化 | 无变化 |
| 阿司匹林 | 无变化 | 无变化 | 延长 | 无变化 |
| 血小板减少症 | 无变化 | 无变化 | 延长 | 减少 |
| 急性肝衰竭 | 延长 | 无变化 | 无变化 | 无变化 |
| 晚期肝衰竭 | 延长 | 延长 | 延长 | 减少 |
| 尿毒症 | 无变化 | 无变化 | 延长 | 无变化 |
| 无纤维蛋白原血症 | 延长 | 延长 | 延长 | 无变化 |
| 缺乏凝血因子Ⅴ | 延长 | 延长 | 无变化 | 无变化 |
| 缺乏凝血因子Ⅹ | 延长 | 延长 | 无变化 | 无变化 |
| 血小板衰弱症 | 无变化 | 无变化 | 延长 | 无变化 |
| 巨大血小板综合症 | 无变化 | 无变化 | 延长 | 减少或无变化 |
| 缺乏凝血因子XII | 无变化 | 延长 | 无变化 | 无变化 |
| 遗传性血管水肿 | 无变化 | 缩短 | 无变化 | 无变化 |
凝固止血物质
●抗凝血酶III (AT3):一种分子量为 65,000 的蛋白质,在肝和血管内皮细胞中表达并释放到血液中。起抑制凝血因子Xa和凝血酶的作用。肝素与变构位点的结合使其活性增加1000倍。
●肝素:一种能激活抗凝血酶III的多糖。低分子量肝素还降低血管性血友病因子的活性并抑制其与血小板的反应。
●蛋白C:被凝血酶分解成活化蛋白 C (APC),与辅酶蛋白 S 结合。它结合并抑制活性凝血因子V和活性凝血因子VIII。
●蛋白S:活性蛋白 C (APC) 的辅酶,与 APC 结合发挥抗凝作用。
●乙二胺四乙酸(EDTA) 和柠檬酸通过螯合血浆中的游离Ca⁺⁺来阻断凝血酶的形成。两者都用于防止采血后的血液凝固。柠檬酸也是人体内的一种成分,其在体内迅速代谢而贷不到影响凝血活性的浓度,因此它也用于体外循环回路和防止输血时储存的血液凝固。另一方面,EDTA不是人体的成分,在体内不代谢,在螯合二价金属离子的同时从尿中排出,发挥抗凝作用后不会返回人体。
●阿司匹林不是人体成分,它是通过抑制环氧合酶和抑制血小板中花生四烯酸合成前列腺素和血栓素A2而发挥抗血小板作用的药物。即使将其直接添加到采集的血液中,也不会阻止凝血。
●华法林是一种用于抑制血栓形成的药物,而不是人体成分。口服华法林,可通过抑制作为Ca结合位点的γ-羧基谷氨酸的产生,阻碍肝脏生物合成凝血因子,从而抑制血液凝固。当添加到被采集的血液中时,它不会阻止凝血。
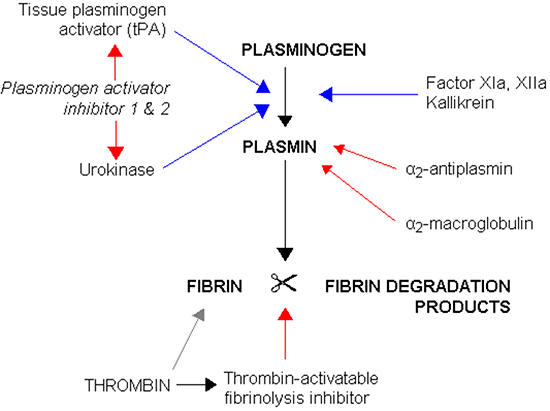
★纤溶系统
凝固的血凝块(血栓)对机体来说是外来物质,在组织修复过程中机体会主动努力清除血栓。纤维溶解系统因此而存在。
●血浆纤溶酶原被组织纤溶酶原激活剂(t-PA)或尿激酶(u-PA)激活,生成纤溶酶。
●纤溶酶将凝固的纤维蛋白分解成D-二聚体和其他分解产物。
纤溶抑制物
●纤溶酶原激活抑制剂
●α1-抗纤溶酶:一种分子量约为51,000的糖蛋白,由位于 14 号染色体长臂末端(14q32)的 SERPINA1 基因编码,有抑制活化纤溶酶的作用。缺乏它会增加患COPD(慢性阻塞性肺病)的风险,但尚不清楚其机制。
●α2-抗纤溶酶:一种分子量为59,000的糖蛋白,由SERPINF1 和 SERPINF2 基因编码,映射到第 17 号染色体 (17p13) 的短臂末端,在肝脏中合成并被释放到血液中。在血浆电泳中属于α2球蛋白,比上述α1抗纤溶酶能更大发挥纤溶抑制作用。
●凝血酶激活的纤溶抑制剂
纤溶异常
正常状态下,血管内不断重复发生血栓形成和纤维蛋白溶解的现象,处于一种平衡状态,周而复始。当失去这种平衡时就会引起各种疾病。
在多发性创伤中,组织因子流入血液激活凝血系统,同时菌血症等内毒素通过诱导炎症介质减弱血管内皮细胞的抗血小板作用,最终导致全身各部位形成血栓。这一过程中凝血因子被消耗,最后可能枯竭。同时,机体为了消除血栓引起的循环系统障碍,纤维蛋白溶解系统被增强,导致无法止血。这就是弥散性血管内凝血(DIC)。肝素用于治疗DIC,但如果AT3耗尽,则无效,因此应该同时给药AT3。另外,由于多数凝血因子和纤溶因子(因子II、VII、IX、XI、XIII和纤溶酶)是由丝氨酸蛋白酶演化而来的,因此应该同时给药丝氨酸蛋白酶抑制剂,例如甲磺酸萘莫司他和甲磺酸加贝酯。
参考文献
Ferguson JJ. et al. "Safe use of platelet GP IIb/IIIa inhibitors." Eur Heart J. 19 Suppl D:D40-51.;1998 Apr EntrzPubMed